Hi Vyntager,
I finally have the opportunity to start CR
Congratulations! What was holding you back before, and how did you overcome it?
I'd have a few questions though :
1 ) Fiddling with programs such as cronometer, ... I almost always end up having like 1000 + % vitamin A (counting beta carotene I suppose), as well as very high vitamin K. I've heard about risks associated with too high a consumption of beta carotene though. Have you already had such issues ? Should I care, or just go on ? Is there something I should change ?
No, you totally don't have to worry about this. Too much
preformed vitamin A (retinol) is really dangerous (increases fracture risk, per a great deal of high-quality prospective epidemiology), but beta-carotene has no such issues; likewise, there are dangers to beta-carotene
supplements, but (a) that's largely in smokers, (b) it probably involved the lack of coantioxidants, © there's a dose effect (ie, I take it you're not getting 25000-50K IU of beta-carotene), and (d) any cancer problem is in part attributable to the fact that most supplements used then, and use now,
synthetic (all-
trans) b-c, which is chemically and 'physiologically' distinct from food-source (
cis-trans) b-c. Similarly, I'd not worry about vitamin K unless you're on warfarin -- and even then, it's fine so long as it's
consistent and your doc is doing proper INR testing.
Additionnally, I have more than 100% proteins, fats, carbs, but not even 100% my recommended Calorie intake. Why is that so ? The best explanation I could think of is it would be because the protein / fat / carbs relates to the minimal amount of any of those you'd need in your regimen, but as I understand the program, that's not the case.
You misunderstand the program

-- that's exactly what it does. The Calorie intake will of course be low: you're on CR! IAC, you can reset your targets.
2 ) I've been reading from different sources (like Walford in the anti-aging plan), that one should start the CR regimen slowly, dropping calories in at least 6-9 months, if not more. The important thing was, if you start CR too abruptly, then you'd loose all the benefits (or alternatively, that loosing too much weigth too fast will make you loose all the benefits of CR).
Could someone point me to the scientifical publications which advanced that idea ? I'm not convinced, it sounds like dogma, and at any rate I wonder why it would be so, and how it has been observed, and deduced it would be the case that starting CR too fast is a bad thing.
Basically, while you seem to be able to get away with a lot with animals initiated at weaning, and while you can still show a lot of short-term metabolic benefits and protection from
acute insults like nasty neurotoxins by throwing animals straight into full-blown CR, every study of
adult-onset CR has failed to extend lifespan. CR was largely considered to be a laboratory curiosity somehow related to development rather than aging (or, more goofily, that aging somehow
was a programmed process proceeding on from development) this reason, and was therfore ignored for five decades. It was Weindruch and Walford who finally proved that you could do it successfully in adult organisms in 1982, and they did it by (a) initiating CR gradually, and (b) being very careful to give the animals the full dose of vitamins, minerals, EFAs, and protein that they gave the AL animals (in weanlings, you can just feed the poor little bastards half of the same chow, which of course leads to them only getting half of all the nutrients). Here is the relevant material from the seminal paper (2):
Rats and mice given restricted diets from about the age of weaning (3 to 6 weeks) show extended mean and maximum survival times (1) and a decreased incidence or delayed onset of several diseases of old age [REFERENCE]. Other strategies for delaying aging in rodents (for example, administration of antioxidants or hormones) … do not cause clear-cut increases in maximum longevities [REFERENCE] or inhibit age-related increases in mortality rates [REFERENCE]. Old rodents that have been subjected to restricted diets since weaning show more youthful physiologic [REFERENCE] and immunologic [REFERENCE] responses than do age-matched controls. Although underfed rodents consume fewer calories than control animals (25 to 50 percent less in most studies), intakes of other essential nutrients (such as vitamins, salts, and protein) may be maintained by diet enrichment [ie, put in more nutrients per Calorie, so that giving them [i]n[/i]% less food yields the same intake of essential nutrients -MR]. "Undernutrition without malnutrition" is the key concept in increasing life-span by dietary restriction.
Little is known about aging processes in rodents subjected to dietary restriction from midway through their usual life-span or later. Such adult-initiated dietary restriction (considered here as starting at or beyond 10 months of age) increases mean survival times (7-11), but the findings on maximum life-spans [and therefore basic aging processes -MR] are less convincing ( [this key footnote is embedded below as a –MR]). Shortened survival has also been observed in rodents on restricted diets since adulthood ([embedded b below]).
We decided that the influences on survival of dietary restriction initiated in adults warranted further study because in previous studies (i) the dietary restriction was imposed abruptly rather than gradually; (ii) the diets were not enriched, so that some of the animals may have been subjected to malnutrition; and (iii) obese rats were often used (14), in which case the effects of the restricted diet may have been due to the inhibition of obesity. We conducted experiments with male mice from two long-lived strains … At 12 to 13 months of age [human equivalent to ~32-38 y –MR] the mice were subjected to gradual dietary restriction on nutrient-enriched diets, [“Control mice (N = 68) were fed enough of diet 1 (~160 kcal/ week) to maintain their initial body weights throughout much of their subsequent life-spans. The mice to be restricted in their diet (N = 67) received diet 2 (~115 kcal/week) for 1 month and diet 3 (-90 kcal/week) thereafter” and] on which they were maintained until they died. [Again, this equates to a very, very gradual imposition in humans – as much as 3 y. –MR] The mean and maximum life-spans of these animals increased by an average of 10 to 20 percent [ie, 2 LS variables in 2 different strains in the study -MR]. Also, spontaneous lymphoma was inhibited by the underfeeding.
a: McCay et al. [REFERENCE] studied slightly underfed 7- to 15-month-old male white rats (which showed 10 percent decreases in body weight). Although these rats lived longer than control rats on normal diets, their maximum survival was ~38 months (not extremely old for rats). Ross [REFERENCE] imposed restricted diets on 10-month-old male rats previously freely fed a commercial diet and found that only one of four semipurified diets improved survival. Continued feeding of the commercial diet but in restricted amounts (=s 60 to 70 percent of the intake of control rats) also increased average life expectancy. Maximum longevities were not reported in this broad study (20 diet groups, N = 10). Ross [REFERENCE]next fed Charles River CD male rats a semipurified diet similar to the one found effective [REFERENCE]. These rats are large (~800 g when given free access to food; ~19, g of food per day for adults). Rats limited at 10 to 12 months to 8 to 10 g of food per day (maintained at body weights of 285 to 350 g) showed lower mortality ratios (that is, life prolonging influence) and greater life expectancies than rats fed ad libitum. Rats restricted to 10 g of food per day when 12 months old could be expected, at 13 months of age, to live an additional 14 months compared to an additional 9 months for controls. More severe dietary restriction (6 g per day) iniposed on 10- to 12- month-old rats was poorly tolerated, with survival shorter than that of unrestricted controls. Stuchlikova et al. [REFERENCE] studied male mice (unstated strain), golden hamsters, and Wistar rats and found that dietary restriction imposed at 12 months of age raised mean life-spans above those of free-fed controls but did not increase maximum life-spans. Mean life-spans in this study were not long (18 to 23 months). Of 400 mice studied, none lived beyond 36 months. Beauchene et al. [REFERENCE] fed Wayne Lab Blox to 12- month-old male Wistar rats, giving them permanent free access to the diet or free access only on alternate nights (no food at other times). The rats on the restricted regimen had a mean lifespan of about 35 months, as opposed to 31 months for controls. Maximum longevities were not reported. … [N]ote that this study involved 25 to 30 rats per group [which is the absolute minimum number required to have a hope of observing a statistically significant effect -MR].
b: BARROWS CH Jr, ROEDER LM.
THE EFFECT OF REDUCED DIETARY INTAKE ON ENZYMATIC ACTIVITIES AND LIFE SPAN OF RATS.
J Gerontol. 1965 Jan;20:69-71. No abstract available.
PMID: 14246534 [PubMed - indexed for MEDLINE]
This result was subsequently duplicated in numerous studies (most of them by Walford and Weindruch, and some by Spindler too (eg, (9-12)), and was absolutely critical to the revival of interest in CR -- and the idea that we could actually
do something about aging -- in the 1980s (as were the human Biosphere results, for which Walford also is responsible). It is somewhat depressing to thinkabout the time that was lost over the previous
five freaking decades after Clive MacKay (18) because no one figured that out earlier.
Unfortunately, many CR studies continue
even now to violate these protocols (and, as a side note,
another important protocol advanced by these investigators, which was to feed the "
ad libitum"
control group
~10-20% less than
literal "ad libitum" (ie, than what they would truly eat if left to their own devices with an unlimited supply of food), in order avoid confounding the effect of CR on basic aging processes with mere obesity-avoidance). Indeed, for a long time the same problem previously seen with
adult-onset CR continued with
late-life CR (after ~16 months): people could show either that you could or could not do a variety of positive-sounding things with late-life, 'shock' CR (eg, (1), but kept trying, and failing, to extend
maximum LS (3-8) -- but when you looked into the study they had violated one or the other or both principles. Finally, Spindler's group at UC Riverside imposed proper, full-nutrition, gradual-onset CR on 19 month old, healthy mice (~54 human years), and got a nice, robust effect, entirely proportional to what was observed in earlier studies with CR imposed at weaning or in young adulthood.
No one has yet ever shown that you can make CR actually retard aging without these 2 key factors (tho' they've not been tested individually).
It seems reasonable to think that the older organism, having an established body layout and already having suffered some homeostasis-impairing aging damage, would be less able to adapt to the shock of suddenly having almost half its food tken away and/or being mildly malnourished.
3 ) I've had this question too, those mice or rats which had been put under CR, have they eventually been reaching a new set point
Yes -- of course. They aren't losing weight continuously for the equivalent of decades of their furry little lives! Here's
one example, from (13); you can also look at the relevant figures in (9-11), tho' in (10) you see a late-life decline associated with terminal aging (just delayed considerably in CR vs AL animals).
would it be possible that for human beings, doing CR would only work for as long as your body hasn't reached a new set point, and afterwards, the effects of CR would diminish ?
No, the effects are definitely proportional to as long as you stay on CR, period; indeed, you can put animals on CR and then put them back on AL later, and they carry the benefit of those months on CR forward, even tho' they become heavier, because they suffered less aging damage during the time they were on CR, leaving them
younger than they would otherwise be (eg (14-16)). Going back on AL just returns them to "normal" aging. See this useful graph from (17):
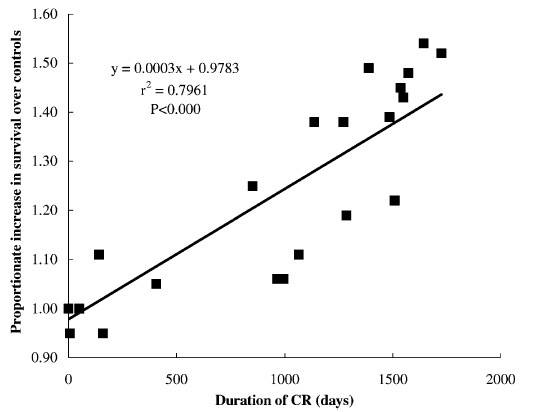
Again, congrats on starting CR!
-Michael
References1. Nagai M, Takahashi R, Goto S. Dietary restriction initiated late in
life can reduce mitochondrial protein carbonyls in rat livers: western
blot studies. Biogerontology. 2000;1(4):321-8. PMID: 11708213
2. Weindruch R, Walford RL.
Dietary restriction in mice beginning at 1 year of age: effect on life-span and spontaneous cancer incidence.
Science. 1982 Mar 12;215(4538):1415-8.
PMID: 7063854 [PubMed - indexed for MEDLINE]
3. Goodrick CL, Ingram DK, Reynolds MA, Freeman JR, Cider NL.
Differential effects of intermittent feeding and voluntary exercise on body weight and lifespan in adult rats.
J Gerontol. 1983 Jan;38(1):36-45.
PMID: 6848584
4. Lipman RD, Smith DE, Blumberg JB, Bronson RT.
Effects of caloric restriction or augmentation in adult rats: longevity and lesion biomarkers of aging.
Aging (Milano). 1998 Dec;10(6):463-70.
PMID: 10078316 [PubMed - indexed for MEDLINE]
5. Lipman RD, Smith DE, Bronson RT, Blumberg J.
Is late-life caloric restriction beneficial?
Aging (Milano). 1995 Apr;7(2):136-9.
PMID: 7548264 [PubMed - indexed for MEDLINE]
6. Forster MJ, Morris P, Sohal RS.
Genotype and age influence the effect of caloric intake on mortality in mice.
FASEB J. 2003 Apr;17(6):690-2. Epub 2003 Feb 5.
PMID: 12586746 [PubMed - indexed for MEDLINE]
7. Merry BJ. Food Restriction and the Aging Process. (Photocopy in my posession; damned if I know what book it came from!).
8. Merry BJ. "Dietary Manipulation of Ageing: An Animal Model." In
Bittles AH & KJ Collins, The Biology of Human Ageing, Cambridge
University Press, Cambridge, 1986.
9. Lee CK, Pugh TD, Klopp RG, Edwards J, Allison DB, Weindruch R, Prolla TA.
Abstract
The impact of alpha-lipoic acid, coenzyme Q10 and caloric restriction on life span and gene expression patterns in mice.
Free Radic Biol Med. 2004 Apr 15;36(8):1043-57.
PMID: 15059645 [PubMed - indexed for MEDLINE]
10. Pugh TD, Oberley TD, Weindruch R.
Dietary intervention at middle age: caloric restriction but not dehydroepiandrosterone sulfate increases lifespan and lifetime cancer incidence in mice.
Cancer Res. 1999 Apr 1;59(7):1642-8.
PMID: 10197641 [PubMed - indexed for MEDLINE]
11. Weindruch R, Gottesman SR, Walford RL.
Modification of age-related immune decline in mice dietarily restricted from or after midadulthood.
Proc Natl Acad Sci U S A. 1982 Feb;79(3):898-902.
PMID: 6977776
12. Spindler SR, Mote PL.
Abstract
Screening candidate longevity therapeutics using gene-expression arrays.
Gerontology. 2007;53(5):306-21. Epub 2007 Jun 15. Review.
PMID: 17570924 [PubMed - indexed for MEDLINE]
[Lifespan curve reproduced (crudely)
here, complete with CR data; CR initiated at 14 months -MR]
12: Dhahbi JM, Kim HJ, Mote PL, Beaver RJ, Spindler SR.
Temporal linkage between the phenotypic and genomic responses to caloric restriction.
Proc Natl Acad Sci U S A. 2004 Apr 13;101(15):5524-9. Epub 2004 Mar 25.
PMID: 15044709 [PubMed - indexed for MEDLINE]
13. Barger JL, Kayo T, Vann JM, Arias EB, Wang J, Hacker TA, Wang Y, Raederstorff D, Morrow JD, Leeuwenburgh C, Allison DB, Saupe KW, Cartee GD, Weindruch R, Prolla TA.
A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice.
PLoS ONE. 2008 Jun 4;3(6):e2264.
PMID: 18523577 [PubMed - in process]
14. Beauchene RE, Bales CW, Bragg CS, Hawkins ST, Mason RL.
Effect of age of initiation of feed restriction on growth, body
composition, and
longevity of rats.
J Gerontol. 1986 Jan;41(1):13-9.
PMID: 3941250 [PubMed - indexed for MEDLINE]
15. Yu BP, Masoro EJ, McMahan CA.
Nutritional influences on aging of Fischer 344 rats: I. Physical,
metabolic, and longevity characteristics.
J Gerontol. 1985 Nov;40(6):657-70.
PMID: 4056321
16. Ross MH.
Length of life and caloric intake.
Am J Clin Nutr. 1972 Aug;25(8):834-8. No abstract available.
PMID: 5046728 [PubMed - indexed for MEDLINE]
17. Merry BJ.
Molecular mechanisms linking calorie restriction and longevity.
Int J Biochem Cell Biol. 2002 Nov;34(11):1340-54. Review.
PMID: 12200030 [PubMed - indexed for MEDLINE]
18. McCay CM, Crowell MF, Maynard LA.
The effect of retarded growth upon the length of life span and upon the ultimate body size.
J Nutr. 1935 Jul 10;10(1):63-79.
Edited by Michael, 20 July 2011 - 08:59 PM.
Replacing missing Merry graph


































 20080705a.jpg 150.39KB
95 downloads
20080705a.jpg 150.39KB
95 downloads

















{kind=link}